Cependant, il manque une taxonomie complète de la latéralisation fonctionnelle et de son organisation dans le cerveau.
Nous rapportons ici la première carte complète des asymétries hémisphériques fonctionnelles dans le cerveau humain, qui révèle sa structure dimensionnelle basse et sa relation avec la connectivité inter-hémisphérique structurelle.
Notre cerveau se compose d’un hémisphère gauche et d’un hémisphère droit remplissant, chacun, des rôles différents. Même si nous utilisons constamment les deux, il est possible, selon sa personnalité, de privilégier les fonctions de l’un ou de l’autre. Alors, plus analytique comme le cerveau gauche ou plus créatif comme le cerveau droit, où vous situez-vous ?
Nos résultats suggèrent que la latéralisation des fonctions cérébrales est répartie selon quatre axes fonctionnels: la communication symbolique, la perception / action, l’émotion et la prise de décision.
La similitude entre cette découverte et les travaux récents sur les symptômes neurologiques donne lieu à de nouvelles hypothèses sur les mécanismes qui soutiennent la récupération du cerveau après une lésion cérébrale.
Nous rapportons également que les régions corticales montrant des asymétries dans l’activité évoquée par la tâche ont des connexions réduites avec l’hémisphère opposé. Ce dernier résultat suggère que, au cours de l’évolution, l’expansion de la taille du cerveau a conduit à une latéralisation fonctionnelle afin d’éviter des délais de conduction excessifs entre les hémisphères.
LA TENTATION DU « DOPAGE » DE CERVEAU
L’objectif est simple : maximiser les gains neurologiques en utilisant la stimulation transcrânienne (ou tDCS).
Ce procédé à base d’électrodes placées dans l’arceau du casque permettrait de stimuler électriquement pendant 20 minutes certains circuits neuronaux les plus utiles à la performance de façon à les rendre plus automatiques. En améliorant la plasticité du cortex cérébral, l’apprentissage serait facilité. On devine facilement le potentiel supposé d’un tel outil au service de sportifs à la recherche de l’infiniment petit qui fera la différence.
Ne dit-on pas que le sportif de haut niveau est un (neuro)chirurgien ? Bien sûr, l’hypothèse n’est viable que si des études suffisamment poussées et élaborées démontrent l’utilité réelle du dispositif, ce qui n’est pas encore le cas aujourd’hui. Pourtant, plusieurs athlètes américains le portent déjà régulièrement. Des équipes cyclistes surveillent aussi cette technologie de près ?—?celle-ci n’étant pas proscrite par l’Agence mondiale antidopage.
INTRODUCTION
La croyance répandue selon laquelle la dominance hémisphérique influence le caractère humain découle d’une interprétation erronée de plusieurs décennies de résultats neuropsychologiques 1 qui montre que la latéralisation fonctionnelle est un principe fondamental de l’organisation du cerveau 2, 3 , 4 . Aujourd’hui, après près de 30 ans de neuroimagerie fonctionnelle, les théories sur la latéralisation fonctionnelle suggèrent une division moins radicale et supposent que les deux hémisphères s’équilibrent 5 . Cependant, malgré les implications des théories de la latéralisation fonctionnelle pour les troubles neurodéveloppementaux et psychiatriques 6 , 7 , ainsi que pour le rétablissement après un AVC 8 ,9 , 10, 11 , une cartographie complète de la latéralisation fonctionnelle dans le cerveau est, à notre connaissance, encore absente de la littérature.
On ne sait pas non plus si les fonctions cognitives supposément latéralisées partagent des patrons spatiaux d’activation fonctionnelle similaires ou différents, et si ces activations fonctionnelles peuvent être classées en un nombre limité de patrons spatiaux – ont une structure de faible dimension.
De plus, les mécanismes qui soutiennent la latéralisation fonctionnelle et la communication inter-hémisphérique associée restent débattus 12 ,13 . Deux hypothèses concurrentes ont été proposées sur l’émergence d’une latéralisation fonctionnelle basée sur la structure du corps calleux, le lien inter-hémisphérique le plus considérable.
L’hypothèse d’indépendance interhémisphérique suggère que, au cours de l’évolution, l’expansion de la taille du cerveau a conduit à une latéralisation fonctionnelle afin d’éviter des délais de conduction excessifs entre les hémisphères 14 . Par conséquent, les régions fonctionnellement latéralisées seront moins connectées via le corpus callosum que les régions non latéralisées afin de rendre le traitement des fonctions latéralisées plus efficace 15. L’hypothèse de la concurrence interhémisphérique suggère que la latéralisation fonctionnelle résulte de la compétition entre les hémisphères qui se inhibent via le corps calleux.
Comme les régions fonctionnellement latéralisées auraient besoin d’inhiber davantage l’hémisphère opposé que les régions non latéralisées, elles pourraient être davantage connectées par le corps calleux. Des études préliminaires anatomiques 16 et d’imagerie par résonance magnétique fonctionnelle (IRMf) 17 appuient les deux théories. Toutefois, le petit nombre de fonctions étudiées et les lacunes des méthodes limitent souvent l’interprétabilité des résultats 13 .
Dans l’ensemble, la généralisation de ces théories et découvertes à l’organisation fonctionnelle du cerveau entier reste inconnue.
Ici, nous avons profité de la combinaison du plus grand ensemble de données méta-analytiques IRMf 18 avec les données de connectivité structurelle de la plus haute qualité 19 pour produire, pour la première fois, une carte complète de l’architecture cérébrale fonctionnelle des fonctions cognitives latéralisées, en caractérisant la structure de faible dimension. et d’examiner sa relation avec la connectivité du corps calleux.
RESULTATS
Cartes de latéralisation fonctionnelle et leur structure en basse dimension
Nous avons sélectionné 590 termes liés à des processus cognitifs spécifiques dans l’ensemble de la base de données Neurosynth (voir le tableau supplémentaire 1 ).
Une carte de latéralisation fonctionnelle a été calculée pour chaque terme en calculant la différence entre les hémisphères de chaque paire de voxels homologues. Régions fonctionnelles homologues peuvent être déplacés dans les deux hémisphères en raison de facteurs anatomiques, par exemple le couple Yakovlevian 4 , 20 , la taille de la planum temporale 21 et le cortex moteur 22 , 23 .
Ici, nous avons ajusté les asymétries anatomiques principales dans les deux hémisphères en enregistrant les cartes dans un atlas symétrique 24 .
Nous avons d’abord cherché à déterminer quelles régions présentaient une latéralisation fonctionnelle significative. Etant donné que les termes sélectionnés pouvaient être corrélés ou liés de manière triviale (par exemple, les formes singulière et plurielle du même mot; « forme visuelle » et « formes visuelles« ), une analyse en composantes principales à rotation varimax a été effectuée afin d’éliminer redondance dans les données.
Cent soixante et onze composantes principales avec des valeurs propres supérieures à la moyenne générale ont été retenues, expliquant 72,6% de la variance des cartes de latéralisation. La modélisation linéaire générale a ensuite été utilisée avec des charges de composants sous la forme d’un ensemble de prédicteurs permettant d’ajuster les données des cartes de latéralisation et d’identifier les voxels associés à une latéralisation significative.
Après 5000 permutations, 25 composants principaux présentaient des voxels avec une latéralisation significative (> 20 voxels;P <0,05 erreur familiale corrigée; voir le tableau supplémentaire 2 ). Celles-ci représentent essentiellement les groupes importants de voxels présentant une latéralisation fonctionnelle significative dans Neurosynth.
Ensuite, une intégration spectrale multivariée, basée sur la similarité entre les cartes de latéralisation, nous a permis d’examiner une structure générique des profils de latéralisation fonctionnelle du cerveau, à savoir son «morphospace» 25 , 26 . L’étape préliminaire, qui incluait l’intégration dans les deux premières dimensions (Fig. 1a et Figure supplémentaire
1 ), a révélé une organisation triangulaire des cartes de latéralisation avec trois sommets: communication symbolique, perception / action et émotion. Un test t -ratio, c’est-à-dire un test d’optimalité de Pareto, 27, entre l’organisation des données réelles et 2000 échantillons de données simulées, obtenus par permutation de l’ordre voxel avant le calcul des différences de gauche à droite, ont confirmé la véracité statistique d’une telle organisation triangulaire. La même analyse a été utilisée pour explorer d’autres dimensions et a révélé trois triangles supplémentaires et un quatrième sommet donné par la prise de décision. (Fig. 1b et Figure supplémentaire 2 ).
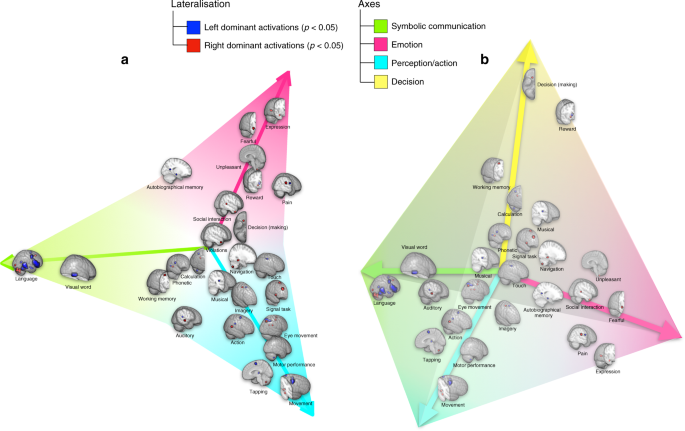
Structure à basse dimension de la latéralisation fonctionnelle. Intégrés dans les espaces bidimensionnels ( a ) et tridimensionnels ( b ) en fonction de la similarité de leur profil de latéralisation (fichier 3D interactif MATLAB disponible en tant que données supplémentaires 1 ). Voir les figures supplémentaires 1 et 2 pour l’intégration spatiale de tous les termes de neurosynth. Ici, afin de fournir un résumé graphique de tous les résultats rapportés dans la section, nous avons tracé les cartes de composants significativement latéralisées, nommées par les termes les plus chargés (Tableau 3 ), à la place des termes réels de Neurosynth (Tableau 1 ).
De plus, en faisant régresser les profils de latéralisation sur les coordonnées des termes dans l’espace intégré, nous avons construit des prédictions pour les cartes situées aux coordonnées des sommets, que nous appellerons cartes d’archétype.
Les cartes archétypes correspondant à l’axe de communication symbolique étaient caractérisées par une activation dominante gauche de la partie postérieure dorsale et ventrale du lobe frontal, y compris la région de Broca et la région motrice présupplementaire, la partie postérieure du lobe temporal, y compris la région de Wernicke et le Zone de formulaire Word visuelle (c.-à-d. VWFA). Les activations dominantes droites étaient localisées dans le lobe postérieur du cervelet, y compris la zone Crus II (Fig. 2a ).

Archétypes de la latéralisation fonctionnelle. Les cartes correspondent à la communication symbolique ( a ), à la perception / action ( b ), à l’émotion ( c ) et à la décision ( d).) les axes. Le panneau supérieur correspond à la vue latérale, le panneau central à la vue médiale et le panneau inférieur à la vue du cervelet (vues latérale et postérieure) du modèle d’activation reconstitué. Zone visuelle sous forme de mots VWFA, zone WA Wernicke, cortex frontal ventral VFC, gyrus frontal inférieur IFg, gyrus frontal moyen MFg, jonction temporo-pariétale en TPJ, lobule pariétal inférieur IPL, cortex squelettique supérieur IP, IP cortex moteur, champ oculaire frontal FEF, lobe temporal postérieur PTL, cortex préfrontal PFC, zone motrice supplémentaire SMA, zone motrice pré-supplémentaire SMA, cortex cingulaire antérieur ACC, cerveau antérieur basal BF (les cartes sont disponibles en tant que Données supplémentaires 2 – 5 )
La carte de perception / action des archétypes impliquait le cortex sensorimoteur gauche, le SMA gauche et le thalamus gauche. Les activations dominantes droites comprenaient le champ oculaire frontal, la région intrapariétale et les régions frontales ventrales, le champ oculaire frontal, la région motrice présupposée, le cerveau antérieur basal et le cervelet antérieur (zones V / VI et VIII) ainsi qu’une partie du vermis (Fig. 2b ). .
La carte des émotions archétypes impliquait le cortex cingulaire antérieur gauche, le complexe basolatéral de l’amygdale droite, la partie postérieure du gyrus frontal inférieur droit, le sulcus intrapariétal droit et la partie postérieure du lobe temporal droit (Fig. 2c ).
Enfin, la carte des archétypes décisionnels impliquait principalement le cortex préfrontal droit (gyrus orbital médial), le champ oculaire frontal droit, le sulcus intrapariétal gauche ainsi que le striatum (putamen droit et caudé gauche) et le cerveau antérieur basal gauche (Fig. 2d ).
Corpus callosum et latéralisation fonctionnelle
Étant donné que la diffusion microscopique des molécules d’eau dans le cerveau est plus facile le long des axones que possible, la tractographie dérivée de l’imagerie par résonance magnétique pondérée en diffusion permet d’analyser l’organisation structurelle de la connectivité cérébrale (Fig. 3a ).
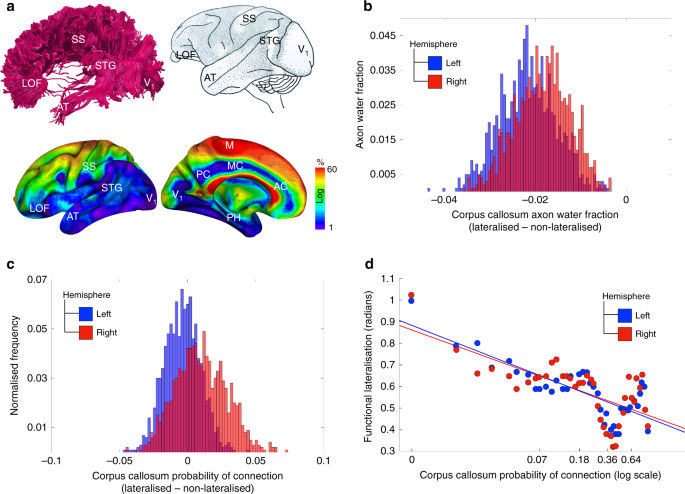
Latéralisation et connectivité interhémisphérique. une tractographie du corps calleux chez un sujet représentatif de notre étude (en haut à gauche); projection corticale du corps calleux dérivée du traçage axonal chez le singe 31 (en haut à droite); projections corticales du corps calleux dérivées de la tractographie chez les participants à notre étude (en bas). bHistogramme de la différence entre les régions latéralisées et non latéralisées de la fraction hydrique axonale du corps calleux, calculée en moyenne pour tous les participants. cHistogramme de la différence entre les régions latéralisées et non latéralisées dans la probabilité de connexion du corps calleux. La mesure a été calculée comme la proportion de participants dans laquelle il existe un lien entre les voxels du cerveau et le corps calleux et la taille de l’échantillon global du professionnel de la santé. d Relation dimensionnelle entre le degré de latéralisation fonctionnelle et la probabilité de connectivité du corps calleux. Cortex orbitofrontal latéral LOF, cortex somatosensoriel SS, gyrus temporal supérieur STG, temporal antérieur AT, aire visuelle primaire V1, aire motrice primaire M, gyrus cingulaire postérieur PC, gyrus cingulaire moyen MC, gyrus cingulaire antérieur, AC
Dans ce qui suit, nous avons analysé la relation entre la latéralisation fonctionnelle et les mesures de connectivité du corps calleux en comparant les profils de connectivité des régions latéralisées et non latéralisées (voir Méthodes de définition des régions non latéralisées). Deux mesures de la connectivité ont été utilisées, les deux étant calculées en faisant la moyenne entre les participants de l’échantillon de HCP. La première mesure, la fraction d’eau axonale 28 , est microscopique et est estimée dans la section médiane du corps calleux traversée par des lignes de courant provenant de voxels d’une région corticale sélectionnée. La deuxième mesure est macroscopique et estime la réplicabilité des connexions 29 , 30 calculée comme la proportion de participants dans laquelle un voxel est connecté au corpus callosum, ce que nous appellerons probabilité de connexion pour raccourcissement.
En échantillonnant des voxels de régions latéralisées et non latéralisées, dans chaque hémisphère séparément, nous avons d’abord construit la distribution des différences dans la fraction d’eau axonale entre les régions latéralisées et non latéralisées. La figure 3b indique que la fraction d’eau axonale était systématiquement plus basse pour les voxels du corps calleux se projetant sur les régions latéralisées par rapport aux voxels non latéralisés. De plus, les graphiques suggéraient une fraction d’eau axonale légèrement inférieure pour les régions de l’hémisphère gauche par rapport à l’hémisphère droit.
Ensuite, nous avons construit une distribution analogue pour la probabilité de connexion. La figure 3c montre que les régions latéralisées comparées aux voxels non latéralisés ne différaient pas dans cette mesure macrostructurale de la connectivité.
L’analyse précédente n’a pas permis de révéler une différence catégorique entre les régions latéralisées et non latéralisées dans la mesure macroscopique de la connectivité. Cependant, le degré de dominance fonctionnelle hémisphérique (voir Méthodes pour la définition de cette mesure) peut varier – d’un modèle d’activité unilatéral à un modèle relativement asymétrique. Dans ce dernier cas, les deux hémisphères sont impliqués dans une fonction, mais l’un est plus actif que l’autre. Nous avons donc cherché à savoir s’il existait une relation proportionnelle entre le degré de latéralisation fonctionnelle et la probabilité de connectivité du corps calleux.
La figure 3d indique une relation négative entre la probabilité de connexion et le degré de latéralisation fonctionnelle, à la fois pour les hémisphères gauche et droit (corrélation de Pearson, r = –0,81 et r = –0,69, respectivement, p <0,001). Comme le niveau global d’activation de deux zones homotopes des hémisphères gauche et droit peut avoir une influence sur les connexions de son corps calleux, nous avons dupliqué la même analyse après avoir régressé le niveau moyen d’activité des hémisphères gauche et droit pour chaque voxel fonctionnellement latéralisé.
La relation entre le niveau de dominance fonctionnelle et la probabilité de connexion au corps calleux est restée inchangée pour l’hémisphère gauche (corrélation de Pearson r = –0,79) et augmenté pour l’hémisphère droit (corrélation de Pearson r = –0,85).
Des analyses complémentaires supplémentaires ont indiqué qu’il n’existait aucune relation entre la différence de connectivité du corps calleux des voxels latéralisés et non latéralisés et leur distance du centre du corps calleux (Figure 3 supplémentaire ).
En conclusion, la présente analyse nous fournit une vue complète de la latéralisation fonctionnelle chez l’homme, qui semble être organisée en quatre domaines: la communication symbolique, la perception / action, les fonctions liées à l’émotion et à la décision. Il révèle également certains de ses mécanismes, tels que la relation entre la latéralisation fonctionnelle et la force de la communication entre les hémisphères. La similitude entre les résultats actuels et les travaux récents sur les symptômes neurologiques donne lieu à de nouvelles hypothèses sur les mécanismes qui soutiennent la récupération du cerveau après une lésion cérébrale.
DISPONIBILITE DES DONNEES
Les données analysées au cours de la présente étude sont disponibles sur les sites https://www.humanconnectome.org et http://www.neurosynth.org .
De plus, les données traitées sont disponibles sur demande aux auteurs correspondants michel.thiebaut?@gmail.com et http: //vyacheslav.karolis?@ndcn.ox.ac.uk.

